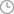1.高山离子芥内质网型ω-3脂肪酸去饱和酶基因在提高烟草抗逆性方面的应用,其特征在于:该高山离子芥内质网型ω-3脂肪酸去饱和酶基因来源于高山离子芥,所述基因编码的内质网型ω-3脂肪酸去饱和酶的氨基酸序列如序列表中SEQ ID NO.2第1位到386位所示,其核苷酸序列如序列表中SEQ ID NO.1第1位到1161位所示。
高山离子芥内质网型ω-3脂肪酸去饱和酶基因及其应用\n技术领域\n[0001] 本发明涉及基因工程技术领域,尤其涉及高山离子芥内质网型ω-3脂肪酸去饱和酶基因及其应用。\n背景技术\n[0002] 低温、干旱和盐渍是自然界常见的非生物胁迫,是影响农作物产量和品质的重要因素。培育能同时抵抗寒冷、干旱和盐胁迫的作物新品种对于提高农业生产力,提高水资源和土地利用率具有重要意义。烟草是一种重要的经济作物,但许多优良品质的烟草自身抗逆性并不强,不仅限制了其种植区域,还使之很容易受到突发气候灾害的影响。因此,鉴定新的抗逆基因,通过基因工程技术提高烟草的综合抗逆性将是改良烟草的重要途径。\n[0003] 脂肪酸去饱和酶(Fatty acid desaturase)是指催化与载体结合的脂肪酸链脱氢形成C=C双键的酶,根据作用底物载体的不同可分为酰基CoA去饱和酶(Acyl-CoA desaturase),酰基ACP去饱和酶(Acyl-ACP desaturase)和酰基脂去饱和酶(Acyl-lipid desaturase)(Los and Murata, 1998)。ω-3脂 肪 酸去 饱 和 酶(ω-3 Fatty acid desaturase)是酰基脂去饱和酶的一种,只存在于植物和蓝藻中,根据其在细胞内的定位可分为“内质网型”和“质体型”(Gibson et al., 1994)。该酶能通过催化与甘油在sn-1或sn-2位置结合的二烯脂肪酸(C16:2或C18:2)在ω-3位形成第三个双键而转化为三烯脂肪酸(C16:3或C18:3)(Ohlrogge and Browse, 1994)。三烯脂肪酸(Trienoic fatty acids, TAs)是高等植物细胞膜中主要的不饱和脂肪酸,它不仅是膜的组成部分,其含量的变化还可响应不同的逆境胁迫,缓解逆境对植物造成的伤害,增强植物的抗逆性(Matsuda et al., 2007)。内质网型ω-3脂肪酸去饱和酶在常温下表达,表达具有组织特异性,并受温度、盐、干旱、光照、伤害和激素等多种因子调节(Somerville and Browse, 1996)。研究表明,内质网型ω-3脂肪酸去饱和酶不仅可通过增加植物三烯脂肪酸含量提高植物抗寒性,对植物在干旱和盐渍等逆境条件下存活也具有重要作用(Shimada et al., 2000; Yu et al., 2009; Zhang et al., 2005; Wang et al., 2013)。这些研究还揭示,通过增加三烯脂肪酸比例(C18:3/C18:2)提高抗逆性的植株在非生物胁迫下能维持较好的膜状态,因此膜损伤程度显著低于普通植株,具有相对更低的膜透性(相对电导率值)和膜脂氧化程度(MDA值)。\n[0004] 高山离子芥(Chorispora bungeana)是十字花科离子芥属多年生草本植物,分布在高海拔亚高山草甸和砾石质山坡上,其生境特点是气候寒冷,土层反复冻融,空气稀薄,辐射强,劲风,且砾石保水性差,常造成干旱胁迫。高山离子芥在外部结构和生态策略上无显著抗性特征,为耐受不利环境,必通过表达抗性基因来维持自身的生理代谢和生长发育,以避免和减轻胁迫造成的危害。因此,高山离子芥是研究植物抗逆性的理想材料。目前,还未见从高山离子芥克隆出内质网型ω-3脂肪酸去饱和酶基因的报道,故从高山离子芥克隆鉴定内质网型ω-3脂肪酸去饱和酶基因,通过基因工程将其转到烟草中,对烟草抗逆性的提高具有重要的现实意义。\n发明内容\n[0005] 本发明所要解决的技术问题是提供一种提高烟草综合抗逆性的高山离子芥内质网型ω-3脂肪酸去饱和酶基因。\n[0006] 本发明所要解决的另一个技术问题是提供基于该基因在提高烟草抗逆性方面的应用。\n[0007] 为解决上述问题,本发明所述的高山离子芥内质网型ω-3脂肪酸去饱和酶基因(Chorispora bungeana fatty acid desaturase 3,CbFAD3),其特征在于:该基因来源于高山离子芥,所述基因编码的内质网型ω-3脂肪酸去饱和酶的氨基酸序列如序列表中SEQ ID NO.2第1位到386位所示。\n[0008] 所述基因具有序列表中SEQ ID NO.2第1位到386位所示的氨基酸残基序列经过一个或多个氨基酸残基的取代、缺失或添加且具有与SEQ ID NO.2的氨基酸残基序列相同活性的由SEQ ID NO.2衍生的蛋白质;所述由SEQ ID NO.2衍生的蛋白质与所述等位基因的序列相对应。\n[0009] 其核苷酸序列如序列表中SEQ ID NO.1第1位到1161位所示。\n[0010] 所述基因具有序列表中SEQ ID NO.1第1位到1161位中因一个或多个碱基的缺失、插入或替换而形成的具有功能活性的等位基因。\n[0011] 一种高山离子芥内质网型ω-3脂肪酸去饱和酶,其特征在于:其氨基酸序列如序列表中SEQ ID NO.2第1位到386位所示。\n[0012] 如上所述的高山离子芥内质网型ω-3脂肪酸去饱和酶基因在提高烟草抗逆性方面的应用。\n[0013] 本发明与现有技术相比具有以下优点:\n[0014] 1、本发明通过核酸序列比对表明,由测序获得的高山离子芥CbFAD3基因编码区全长1161bp如SEQ ID NO.1所示,与模式植物拟南芥的AtFAD3基因同源性最高,达91%。\n本发明通过分别测定高山离子芥CbFAD3基因和拟南芥 AtFAD3基因在低温、干旱和盐胁迫下的表达变化发现:4℃低温下,高山离子芥CbFAD3基因的表达量是25℃下的7.3倍,在胁迫后3小时就达到该峰值;而拟南芥的AtFAD3基因只增加了3.7倍,在胁迫后24小时才达到该值。此外,高山离子芥CbFAD3基因在干旱和盐胁迫下的增加量和响应时间也都优于拟南芥AtFAD3基因。从而表明本发明高山离子芥的 CbFAD3基因对上述胁迫更加敏感,可在较短时间内对胁迫作出响应,且响应的力度更大,故具有更高的抗逆性。\n[0015] 2、本发明通过氨基酸序列分析发现,由SEQ ID NO.1推导出的如SEQ ID NO.2所示的氨基酸序列编码386个氨基酸,其分子量为44.18KD。该氨基酸序列包含高等植物ω-3脂肪酸去饱和酶基因所特有的三个保守组氨酸簇“HDCGH”,“ HGWRISHRTHH”和“HHDIGTHVIHH”,因此属于ω-3脂肪酸去饱和酶家族成员。\n[0016] 本发明通过将高山离子芥的CbFAD3基因连接到载体pYes2上导入酿酒酵母菌INVScl中进行半乳糖诱导的真核表达,表达蛋白大小与SEQ ID NO.2所示的氨基酸序列相符(如图1所示),并在加入外源亚油酸C18:2后催化该转基因酵母产生出了亚麻酸C18:3(如图2所示),从而验证了:如SEQ ID NO.1第1位到1161位的基因序列能表达出具有功能活性的如SEQ ID NO.2中第1位到386位所示的氨基酸残基序列的蛋白质。\n[0017] 3、本发明通过将高山离子芥的CbFAD3基因连接到载体pBI121上导入农杆菌GV3101中再转入烟草本塞姆氏(Nicotiana benthamiana)。利用气相色谱-质谱联用仪对转基因植株的总脂肪酸进行分析发现,其根、茎、叶的C18:3/C18:2比例分别是野生型对照烟草的3.1倍、1.6倍和1.9倍(如图9所示),表明转入的高山离子芥CbFAD3基因的确显著提高了烟草的三烯酸含量。通过低温、干旱和盐胁迫实验发现,转基因烟草的生长状态(如图3~8所示)和膜状态的生理指标(如图10所示)均明显好于野生型对照烟草。这些均说明高山离子芥CbFAD3基因确实能同时提高转基因烟草的抗寒、抗旱和抗盐能力。\n[0018] 4、由于本发明中的高山离子芥的CbFAD3基因具有较强的产生三烯酸的能力和抗逆性,因此,利用此基因通过基因工程技术可培育具有综合抗逆性的烟草,从而改良烟草的品种。\n附图说明\n[0019] 下面结合附图对本发明的具体实施方式作进一步详细的说明。\n[0020] 图1为本发明高山离子芥CbFAD3基因在酿酒酵母INVScl中由半乳糖诱导3d的真核表达蛋白在SDS-PAGE胶上的电泳分析。\n[0021] 图2为本发明含高山离子芥CbFAD3基因的酿酒酵母INVScl在加入亚油酸C18:2后的总脂肪酸气相色谱-质谱联用仪图谱。其中A为加入半乳糖诱导前;B为加入半乳糖诱导后。\n[0022] 图3为本发明转高山离子芥CbFAD3基因的烟草L2和野生型对照在25℃的生长状态。其中:WT为野生型对照;L2为转基因植株。\n[0023] 图4为本发明转高山离子芥CbFAD3基因的烟草L2和野生型对照在-2℃处理24hr并在4℃恢复4hr后的生长状态。其中:WT为野生型对照;L2为转基因植株。\n[0024] 图5为本发明转高山离子芥CbFAD3基因的烟草L2和野生型对照在正常浇水时的生长状态。其中:WT为野生型对照;L2为转基因植株。\n[0025] 图6为本发明转高山离子芥CbFAD3基因的烟草L2和野生型对照在断水10d并复水4hr后的生长状态。其中:WT为野生型对照;L2为转基因植株。\n[0026] 图7为本发明转高山离子芥CbFAD3基因的烟草L2和野生型对照在正常培养基下的生长状态。其中:WT为野生型对照;L2为转基因植株。\n[0027] 图8为本发明转高山离子芥CbFAD3基因的烟草L2和野生型对照在施加10%NaCl处理24hr后的生长状态。其中:WT为野生型对照;L2为转基因植株。\n[0028] 图9为本发明转高山离子芥CbFAD3基因的烟草L2和野生型对照的不饱和脂肪酸比例变化。其中:WT为野生型对照;L2为转基因植株。\n[0029] 图10为本发明转高山离子芥CbFAD3基因的烟草L2和野生型对照在低温、干旱和盐胁迫处理前后的相对电导率变化。其中:WT为野生型对照;L2为转基因植株。\n具体实施方式\n[0030] 在本发明的下列实施例中,所用的实验材料为高山离子芥(Chorispora bungeana)和烟草本塞姆氏(Nicotiana benthamiana) 。\n[0031] 实施例1 高山离子芥内质网型ω-3脂肪酸去饱和酶基因,该基因编码的内质网型ω-3脂肪酸去饱和酶的氨基酸序列如序列表中SEQ ID NO.2第1位到386位所示,其核苷酸序列如序列表中SEQ ID NO.1第1位到1161位所示。\n[0032] 一、高山离子芥CbFAD3基因克隆。\n[0033] 1. 高山离子芥CbFAD3基因保守序列扩增:\n[0034] 1.1. 利用Omega 公司的Plant RNA 提取试剂盒提取高山离子芥愈伤组织中的总RNA。\n[0035] 1.2. 利用Takara公司的PrimeScript™ 1st Strand cDNA合成试剂盒进行反转录反应获得cDNA。\n[0036] 1.3. 根据美国NCBI(National Center forBiotechnology Information)数据库检索的基因序列设计以下简并引物:\n[0037] Sense Primer: 5’-GGGCGGCKATTCCTAAGCAYT-3’(简并度=4)\n[0038] Antisense Primer: 5’-GCRAGCATGGGSAGAGGRACAG-3’ (简并度=8)[0039] 1.3. 高山离子芥CbFAD3基因保守序列PCR扩增:\n[0040] 反应体系(50μl):cDNA 2.5μl,TaKaRa LA Taq(5 U/μl) 0.5 μl,10×LA Taq Buffer II(Mg2+ free)5μl, MgCl2(25 mM)5μl,dNTP Mix(2.5 mM) 8μl,Sense Primer 1μl (10μM),Antisense Primer 1μl (10μM),ddH2O 27μl。\n[0041] 反应条件:94℃,3min\n[0042] (94℃,1min;56℃,15s;61℃,15s;72℃,1min)30个循环\n[0043] 72℃,10min\n[0044] 1.4. 高山离子芥CbFAD3基因保守序列测序:\n[0045] ⑴ 将上述PCR产物在1%的琼脂糖凝胶上进行电泳(220V,170A),在紫外灯下切下符合扩增大小的条带,用Axygen 公司的DNA凝胶回收试剂盒回收。\n[0046] ⑵高山离子芥CbFAD3基因保守序列的克隆和阳性重组子的鉴定按照TransGen公司的pEASY-Blunt Cloning试剂盒进行。\n[0047] ⑶将鉴定出的阳性重组子送至上海Majorbio公司测序。\n[0048] 2. 高山离子芥CbFAD3基因3’端扩增:\n[0049] 2.1. 根据高山离子芥CbFAD3基因保守序列设计以下特异性引物:\n[0050] Outer Primer: 5’- TGAAAACGACGAGTCCTGGGTTCCGCTA-3’\n[0051] Inner Primer: 5’- CTTCACTCCTTCATCCTCGTTCCTTACC -3’\n[0052] 2.2. 高山离子芥CbFAD3基因cDNA(3’RACE)的合成根据Invitrogen公司的SmartRACE试剂盒说明书进行。\n[0053] 2. 3.根据Invitrogen的SmartRACE试剂盒进行PCR 反应:\n[0054] ⑴ Outer PCR 反应。\n[0055] 反应体系(50μl):cDNA(3’RACE) 2.5μl,Outer primer (10uM) 1μl,10×UPM \n5μl,ddH20 34.5μl,10×BD Advantage 2 PCR Buffer 5μl,dNTP Mix(20mM)1μl,\n50×BD Advantage 2 Phymerase Mix 1μl。\n[0056] 反应条件:(94℃ 30s;68℃,30s;72℃,3min)25个循环。\n[0057] ⑵ Inner PCR 反应。\n[0058] 反应体系(50μl):Outer PCR稀释产物 5μl,Inner primer (10uM) 1μl,\n10×NUP 5μl,ddH20 32μl,10×BD Advantage 2 PCR Buffer 5μl,dNTP Mix(20mM)\n1μl,50×BD Advantage 2 Phymerase Mix 1μl。\n[0059] 反应条件同本实施例中2.3⑴。\n[0060] 2.4. 高山离子芥CbFAD3基因3’端测序方法同本实施例中1.4。\n[0061] 3. 高山离子芥CbFAD3基因5’端扩增:\n[0062] 3.1.根据高山离子芥CbFAD3基因保守序列设计以下特异性引物:\n[0063] Outer Primer: 5’- GAGGCACAGTGTATCTGAGCATTCGA -3’\n[0064] Inner Primer:5’- GGTAAGGAACGAGGATGAAGGAGTGAAG -3’\n[0065] 3.2. 高山离子芥CbFAD3基因cDNA(5’RACE)的合成根据Invitrogen公司的SmartRACE试剂盒说明书进行。\n[0066] 3. 3.根据Invitrogen的SmartRACE试剂盒进行PCR 反应:\n[0067] ⑴ Outer PCR 反应。\n[0068] 反应体系(50μl):cDNA(5’RACE) 2.5μl,其余反应剂同本实施例中2.3⑴。\n[0069] 反应条件同本实施例中2.3⑴。\n[0070] (2) Inner PCR 反应。\n[0071] 反应体系(50μl)和反应条件同本实施例中2.3⑴。\n[0072] 3.4. 高山离子芥CbFAD3基因5’端测序方法同本实施例中1.4。\n[0073] 4. 基因拼接:\n[0074] 4.1. 将高山离子芥CbFAD3基因保守序列,3’ 端和5’ 端基因测序结果通过DNAstar软件拼接,得到基因cDNA序列全长。\n[0075] 5. 基因全长cDNA 序列扩增:\n[0076] 5.1. 利用cDNA 为模板设计引物如下:\n[0077] CbFAD3s:5’-GCTCTAGAATGGTTGTTGCAATGGACCA-3’\n[0078] CbFAD3a:5’-CGAGCTCCTAATTGATTTTAGATTTGTCAG-3’\n[0079] 反应体系(50μl):cDNA 2.5μl,TaKaRa LA Taq(5 U/μl)0.5 μl,10×LA Taq Buffer II(Mg2+ free)5 μl,MgCl2(25 mM)5 μl,dNTP Mix(2.5 mM)8 μl,CbFAD3s(10μM) 1μl,CbFAD3a(10μM) 1μl,ddH2O 27μl。\n[0080] 反应条件:94℃,3min\n[0081] (94℃,1min;58℃,30s;72℃,1min)30个循环\n[0082] 72℃,10min\n[0083] 5.2. 高山离子芥CbFAD3基因cDNA序列测序方法同本实施例中1.4。测序得到的CbFAD3基因的编码区如序列表中SEQ ID NO.1所示,自起始密码子至终止密码子总长1161个碱基。\n[0084] 实施例2 高山离子芥CbFAD3基因在酿酒酵母INVScl中的真核表达[0085] 1. 构建真核表达载体pYES2- CbFAD3\n[0086] 1.1.根据编码区序列设计引物如下:\n[0087] Z3-UP:5’-GGATCCACCATGGTTGTTGCAATGGACCA -3’\n[0088] Z3-DN:5’- CGAGCTCTAATTGATTTTAGATTTGTCAG -3’\n[0089] 1.2. 高山离子芥CbFAD3基因扩增:\n[0090] 反应体系(50μl):cDNA 2.5μl,TaKaRa LA Taq(5 U/μl)0.5 μl,10×LA Taq Buffer II(Mg2+ free)5 μl,MgCl2(25 mM)5 μl,dNTP Mix(2.5 mM)8 μl,CbFAD3s(10μM) 1μl,CbFAD3a(10μM) 1μl,ddH2O 27μl。\n[0091] 反应条件:94℃,3min\n[0092] (94℃,1min;56℃,15s;63℃,15s;72℃,1min)30个循环\n[0093] 72℃,10min\n[0094] 1.3. 高山离子芥CbFAD3基因的回收方法同实施例1中1.4.⑴。\n[0095] 1.4.高山离子芥CbFAD3基因与表达载体pYES2质粒的酶切和连接:\n[0096] ⑴将回收产物和表达载体pYES2质粒(FunGenome公司)用Takara公司的限制性内切酶BamHI和SacI分别进行双酶切。反应体系(20μl) :DNA 7μl,BamHI(50 U/μl) 1 μl,SacI1(50 U/μl) 1μl ,10×K Buffer 1 μl,ddH2O 10μl。37℃,酶切4hr。\n[0097] ⑵酶切产物的回收方法同实施例1中1.4.⑴。\n[0098] ⑶将酶切回收后的待插入片段与载体按照Takara公司的DNA Ligation 试剂盒进行连接。\n[0099] ⑷取5μl连接产物加入50μl大肠杆菌DH5α的感受态细胞中进行转化,采用氨苄青霉素(Amp)抗性筛选,长出的菌落直接用PCR鉴定,将鉴定出的阳性克隆送至上海Majorbio公司测序,测序结果得到的CbFAD3基因cDNA序列的编码区如序列表SEQ ID NO.1所示,证实表达载体pYES- CbFAD3构建成功。\n[0100] 2. 表达载体pYES- CbFAD3导入酿酒酵母INVScl\n[0101] 2.1. 利用Axygen公司的质粒抽提试剂盒提取含有表达载体pYES- CbFAD3的阳性克隆的质粒。\n[0102] 2.2. 将含有表达载体pYES- CbFAD3的质粒按照FunGenome公司的高效酵母转化试剂盒导入酿酒酵母INVScl中。\n[0103] 2.3. 利用FunGenome公司的高效酵母质粒回收试剂盒回收酵母质粒,并送至上海Majorbio公司测序,测序结果得到的CbFAD3基因的编码区如序列表SEQ ID NO.1所示,证实表达载体pYES- CbFAD3已成功导入酿酒酵母INVScl中。\n[0104] 3. 蛋白的表达和鉴定:\n[0105] 3.1. 表达蛋白的提取:\n[0106] ⑴将含有表达载体pYES- CbFAD3的酿酒酵母INVScl用专用培养液(0.3%YSDMS+\n0.26%YNB+0.75%(NH4)2SO4+1%NP-40+2%棉籽糖)在28℃,180rpm摇培48hr。诱导时加入2%半乳糖培养3d。\n[0107] ⑵诱导培养结束后,分别吸取1ml未诱导表达和已诱导表达的菌液于1.5ml的离心管中,5000rpm离心3min,弃上清。加入1ml ddH2O重悬后,5000rpm离心3min,弃上清。\n重复操作一次。加入100μl 2×SDS-PAGE上样缓冲液(100mmol/L Tris pH6.8,4%SDS,0.2%溴酚蓝,20%甘油,2%β巯基乙醇),100℃水浴10min。冷却至室温,3000rpm,离心10s。取\n5μl上清液进行SDS-PAGE电泳(稳压:浓缩胶80V,分离胶120V)。诱导表达出的蛋白大小约为44.18KD(如图1所示),与序列表SEQ ID NO.2所示的氨基酸序列预测相符。\n[0108] 3.2. 表达蛋白的功能验证:\n[0109] ⑴含有表达载体pYES- CbFAD3的酿酒酵母INVScl的培养液和培养方法同本实施例中3.1.⑴。诱导时加入2%半乳糖和终浓度为100μM的C18:2(购自sigma公司)培养\n3d。\n[0110] ⑵诱导培养结束后,分别收集20ml未诱导表达和已诱导表达的菌液于50ml的离心管中,5000rpm离心3min,弃上清。加入20ml ddH2O重悬后,5000rpm离心3min,弃上清。\n重复操作一次。加入5ml 1M NaOH(甲醇),70℃水浴30min。冷却后加入6M HCl调pH值至1。加入5ml 10%BF3(甲醇),70℃水浴5min。冷却后加入10ml 饱和NaCl溶液,以5ml CH2Cl2/正己烷(1:4,V:V)萃取两次,合并萃取液干燥得总脂肪酸。\n[0111] ⑶总脂肪酸成分分析使用美国Agilent 公司6890N 型气相色谱和5975C 质谱联用仪。\n[0112] 色谱条件:石英毛细管柱( Agilent DB-FFAP,30.0m×250.0μm×0.5μm),进样量为0.2μl;分流比为100:1;进样口温度为200℃;程序升温从70℃起,以10℃/min升至\n190℃,保持2min,再以8℃/ min升至230℃保持12 min;柱流速为1.1 mL/min;载气为高纯氦气(99.999% )。\n[0113] 质谱条件:电离源为70 eV;离子源温度230℃;四级杆温度150℃;界面温度为\n250℃; 溶剂延迟为1. 5 min。\n[0114] 结果显示(如图2所示)含有表达载体pYES- CbFAD3的酿酒酵母INVScl能催化产生的亚麻酸C18:3。说明如序列表SEQ ID NO.1所示的基因编码区的确能表达出有催化亚油酸C18:2脱氢产生亚麻酸C18:3活性的如序列表SEQ ID NO.2的氨基酸残基所示的蛋白质。\n[0115] 实施例3 转高山离子芥CbFAD3基因烟草的获得\n[0116] 1.构建真核表达载体pBI121-CbFAD3\n[0117] 1.1. 根据编码区序列设计引物如下:\n[0118] ZF3:5’-GCTCTAGAATGGTTGTTGCAATGGACCA- 3’\n[0119] ZF3’:5’ -CGAGCTCCTAATTGATTTTAGATTTGTCAG- 3’\n[0120] 1.2. 高山离子芥CbFAD3基因扩增:\n[0121] 反应体系同实施例2中1.2\n[0122] 反应条件:94℃,3min\n[0123] (94℃,1min;60℃,30s;72℃,1min)30个循环\n[0124] 72℃,10min\n[0125] 1.3.高山离子芥CbFAD3基因的回收方法同实施例1中1.4.⑴。\n[0126] 1.4.高山离子芥CbFAD3基因与表达载体pBI121质粒的酶切和连接:\n[0127] ⑴将1.2的回收产物和表达载体pBI121质粒(本实验室保存)用Takara公司的限制性内切酶XbaI和SacI分别进行双酶切。反应体系(20μl) :DNA 7μl,XbaI(50 U/μl) \n1 μl,SacI1(50 U/μl) 1μl,10×M Buffer 2 μl,ddH2O 10μl。37℃,酶切4hr。\n[0128] ⑵酶切产物的回收方法同实施例1中1.4.⑴。\n[0129] ⑶将酶切回收后的待插入片段与载体按照Takara公司的DNA Ligation 试剂盒进行连接。\n[0130] ⑷取5μl连接产物加入50μl大肠杆菌DH5α的感受态细胞中进行转化,采用卡那霉素(Kan)抗性筛选,长出的菌落直接用PCR鉴定,将鉴定出的阳性克隆送至上海Majorbio公司测序,测序结果得到的CbFAD3基因cDNA序列的编码区如序列表SEQ ID NO.1所示,证实表达载体pBI121-CbFAD3构建成功。\n[0131] 2. 表达载体pBI121-CbFAD3导入农杆菌GV3101\n[0132] 2.1.利用三亲交配法将构建的质粒pBI121-CbFAD3从大肠杆菌DH5α转移到农杆菌GV3101。\n[0133] 2.2.利用Axygen公司的质粒抽提试剂盒提取含有表达载体pYES- CbFAD3的阳性克隆的质粒,并送至上海Majorbio公司测序,测序结果得到的CbFAD3基因的编码区如序列表SEQ ID NO.1所示,证实表达载体pBI121-CbFAD3已成功导入农杆菌GV3101中。\n[0134] 3.利用叶盘法转化烟草:\n[0135] 挑取含有pBI121-CbFAD3的单菌落,接种到1ml YEP 培养液中(Rif 40μg/ml,Kan 100μg/ml) 于28 ℃,180rpm培养约12hr至OD 600为0.6~0.8。将菌液按1%的比例,转入50ml新的含抗生素的YEP培养液中,于28 ℃,180rpm培养6hr至OD600为\n2\n0.4~0.6。取野生型烟草无菌苗的幼嫩叶片,去主脉,剪成0.5cm的小块,放入菌液中,浸泡\n10min。用无菌滤纸吸去多余的菌液后,将叶片接种在MS+KT(2.0μg/ml)+NAA(0.4μg/ml) 固体培养基上,28℃暗培养2d。将共培养后的叶片转移到MS+KT(2.0μg/ml)+NAA(0.4μg/ml)+Kan(80μg/ml)+Cef(300μg/ml) 固体培养基上进行筛选培养,12h/d光照培养,诱导芽的生长。30d后,待不定芽长到1cm 左右时,切下不定芽并转移到1/2MS+NAA(0.1μg/ml) +Cef(200μg/ml) 上进行生根培养,获得完整植株。\n[0136] 4. 转基因植株验证:\n[0137] 利用Omega公司的磁珠植物DNA提取试剂盒提取转化烟草的总DNA,PCR 检测CbFAD3基因是否转入,对扩增出合适条带的植株进一步提取总RNA(提取方法同实施例1中\n1.1),获得cDNA,以cDNA 为模板进一步进行PCR 检测,鉴定转化成功的转基因植株。结果发现5株成功的转基因植株,分别命名为L1,L2,L3,L4和L5。\n[0138] 实施例4高山离子芥内质网型ω-3脂肪酸去饱和酶基因在提高烟草抗逆性方面的应用。通过对转基因烟草的总脂肪酸分析和抗逆性检测表明转基因烟草的抗寒、抗旱和抗盐能力均显著高于野生型对照。\n[0139] 1. 转基因烟草的总脂肪酸分析:\n[0140] 选取经MS+KT(2.0μg/ml)培养基筛选过的转基因植株L2的F1代(仍命名为L2)作为总脂肪酸分析对象。分别取1g待检测株的根、茎、叶,在研钵中加石英砂研磨。总脂肪酸提取和分析方法同实施例2中3.2.⑵~⑶。结果显示,转 CbFAD3基因烟草根、茎、叶的C18:3/C18:2比例均高于野生型对照烟草(如图9所示),表明转入的高山离子芥CbFAD3基因的确显著提高了烟草的三烯酸含量。\n[0141] 2.转基因烟草的抗逆性检测:\n[0142] 选取经MS+KT(2.0μg/ml)培养基筛选过的转基因植株L2的F1代(仍命名为L2)作为抗寒、抗旱和抗盐性检测的对象,将野生型作为对照植株。低温胁迫:将转基因植株与对照植株同时置于-2℃处理24hr后,再于4℃恢复4hr。干旱胁迫:将转基因植株与对照植株同时自然干旱10day,然后再复水4hr。盐胁迫:同时给转基因植株与对照植株施加\n10%NaCl溶液处理24hr。在上述处理完成后立即观察植株生长状态,并测定叶片质膜例子渗透率。结果发现,野生型在处理后植株受损严重,叶片明显萎焉,而转基因植株尚保持较好的生活力(如图3~8所示),转基因植株的质膜渗漏明显小于野生型(如图10所示)。表明转基因烟草的抗寒、抗旱和抗盐能力均显著高于野生型对照。
法律信息
- 2020-09-08
专利权的转移
登记生效日: 2020.08.20
专利权人由中国科学院寒区旱区环境与工程研究所变更为中国科学院西北生态环境资源研究院
地址由730000 甘肃省兰州市城关区东岗西路320号变更为730000 甘肃省兰州市城关区东岗西路318号
- 2015-09-23
- 2014-05-21
实质审查的生效
IPC(主分类): C12N 15/53
专利申请号: 201410024462.7
申请日: 2014.01.20
- 2014-04-23
引用专利(该专利引用了哪些专利)
序号 | 公开(公告)号 | 公开(公告)日 | 申请日 | 专利名称 | 申请人 | 该专利没有引用任何外部专利数据! |
被引用专利(该专利被哪些专利引用)
序号 | 公开(公告)号 | 公开(公告)日 | 申请日 | 专利名称 | 申请人 | 该专利没有被任何外部专利所引用! |