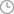1.一种提高植物有用次生物质含量的基因工程方法,其特征是包括步骤:a)构建嵌合基因,包括编码在植物细胞中起作用的启动子的核酸片段,和与启动子为反向连接,编码植物木质素合成关键酶羟基肉桂酰辅酶A还原酶CCR的核酸片段,及转录终止区;b)将嵌合基因导入当其为非转化植物时,即能在其器官或组织中产生积累有用次生物质的选自大豆或红豆杉的植物细胞中,产生转基因植物细胞;c)将转基因植物细胞生长在适宜嵌合基因表达的条件下;d)筛选与鉴别转基因植物细胞,与同类非转基因植物细胞相比,其木质素合成关键酶CCR和木质素含量被减少,而至少有一种有用次生物质的含量被提高;e)将转基因植物细胞进一步培养成为转基因植株,或转基因愈伤组织,或转基因细胞系。
2.根据权利要求1所述的方法,其特征是所述有用次生物质为由苯丙烷途径提供前体的产物。
3.根据权利要求1所述的方法,其特征是所述有用次生物质为药用植物有效成份。
4.根据权利要求1所述的方法,其特征是所述有用次生物质为异黄酮或紫杉醇。
5.一种由权利要求1所述方法获得的有用次生物质含量被提高的转基因植物细胞,其特征是含有所述构建的嵌合基因。
6.一种由权利要求1所述方法获得的有用次生物质含量被提高的转基因植物细胞系,其特征是由所述转基因植物细胞进一步培养形成。
7.一种由权利要求1所述方法获得的有用次生物质含量被提高的转基因植物愈伤组织,其特征是由所述转基因植物细胞进一步培养形成。
一种提高植物有用次生物质含量的基因工程方法\n技术领域\n本发明涉及植物育种技术领域,尤其是关于一种利用基因工程技术提高植物有用次生代谢产物的方法。\n背景技术\n植物次生代谢产生许多有实用价值的天然产物,是很多药物,化妆品,香料,色素等的原料。传统药材含有的有效成份大部分是次生代谢产物,但在天然植物中,这些有用次生物质的含量通常就很低,在栽培和组培植物中更低。因此提高药用植物有效成份含量,是有效利用植物资源,保证中药质量和中药资源可持续利用所迫切需要解决的问题。木质素是植物体中总量仅次于纤维素的第二大有机物质,占植物次生物质总量的95%以上(Anterola A,Lewis N.Trends in Ligninmodification:a comprehensive analysis of the effects of geneticmanipulations/mutations on lignification and vascular integrity.Phytochemistry.2002,61(3):221~294)。木质素及其它许多重要次生物质如植物药的重要成份黄酮类、木脂素、酚类、生物碱、花色素等均由苯丙烷途径(Phenylpropanoid Pathway)合成(http://www.genome.ad.jp/kegg/pathway/map/map00940.html),它们对共同前体(底物)存在着竞争性利用关系。木质素虽在植物体内具有一定的生理功能,但其过多地合成既消耗了大量的碳架结构和能量,也是造纸工业的污染源,并增加其生产能耗;在饲草中木质素含量过高会降低其消化吸收率。所以近年来人们已较重视木质素含量调控,已成为植物基因工程研究的一个重要方向。\n羟基肉桂酰辅酶A还原酶(cinnamoyl CoA reductase,CCR)可还原3种羟基肉桂酸的COA酯,生成相应的肉桂醛(Pichon M,courbou I,Beckert M et al.Cloning and characterization of two maize cDNAsencoding cinnamoyl-CoA reductase(CCR)and differential expression ofthe corresponding genes.Plant Mol.Bio.1998,38(4):671~676)。该酶催化木质素单体生物合成途径的第一个特异步骤,因此对进入木质素合成途径的碳流具有调控作用。Abbott等(Abbott JC,Barakate A,PinconG et al.Simultaneous suppression of multipe genes by singletransgenes.Down-regulation of three unrelated lignin biosynthetic genesin tobacco.Plant Physiol.2002,128:844~853)在烟草上的研究证明了抑制CCR基因表达可大幅度降低植株木质素含量。这些研究结果显示了基因工程技术调控CCR基因表达来改变植物木质素含量的可能。此外,目前已在拟南芥、烟草、扬树等多种植物上获得了木质素含量或成份改变的转基因植株。已证明对木质素含量具有控制作用的关键酶基因包括:PAL(L-phenylalanine ammonia-lyase,苯丙氨酸解氨酶)(Bate N,Orr J,Ni W.et a;.Quantitytive relationship betweenphenylalanine ammonia-lyase levels and phenylpropanoid accumulationin transgenic tobacoo identifies a rate-determining step in natural productsynthesis.1994,PNAS,91:7608-7612),C4H(cinnamate 4-hydroxylase,肉桂酸-4-羟化酶)(Blee K,Choi JW,O’Connell AP et al.Antisense andsense expression of cDNA coding for CYP73A15,a class II cinnamate4-hydroxylase,leads to a delayed and reduced production of lignin intobacco.Phytochemistry 2001,57(7):1159-66),4CL(4-coumarate:coenzyme A ligase,羟基肉桂酸:辅酶A连接酶)(Kajia S,KatayamaY,Omori S.Alteration in the biosynthesis of ligninin transgenic plants with chimeric genes for 4-coumarate:coenzyme Aligase.Plant Cell Physiol.1996,37:957-965),COMT(caffeic acidO-methyltransferase,咖啡酸甲基转移酶)(Atanassova R,Favet N,Martz F,Chabbert b,Tollier M-T,Monties B,Fritig B,Legrand M(1995)Altered lignin composition in transgenic tobacco expressingO-methyltransferase sequences in sense and antisense orientation.Plant J8:465-477),CCoAOMT(caffeoyl-CoA 3-O-methyltransferase,5-腺苷-甲硫氨酸:咖啡酰辅酶A/5-羟基阿魏酰辅酶A-O-甲基转移酰)(ZhongR,Morrison WH,Himmelsbach DS et al.Essential role of caffeoylcoenzyme A O-methyltransferase in lignin biosynthesis in woody poplarplants.Plant Physiol.2000,124:563-577),CCR(cinnamony-CoAreductase,羟基肉桂酰辅酶还原酶)。通过抑制这些酶基因的表达,均可降低植物木质素含量。\n但有关木质素基因工程研究,目前主要集中于植物木质素含量的降低和其组分的改变,而尚未见木质素含量的调控对其它次生代谢途径产物合成影响的系统研究报道;此外,在细胞和组织培养中,采用抑制剂降低竞争途径代谢流来提高目标产物含量已有成功,但通过抑制竞争途径关键酶基因表达来为所需次生产物合成途径提供更多底物的研究尚未见报道。本发明利用“底物竞争”机制,采用基因工程技术调节有关木质素合成特异酶系CCR基因表达,降低木质素生物合成量,为药用植物活性成分等有用次生物质合成提供更多底物,从而提高植物有用次生物质含量。\n发明内容\n本发明的目的是提供一种提高植物有用次生物质含量的基因工程方法,即应用基因工程技术,调节CCR木质素合成关键酶基因表达,降低木质素生物含量,为其它有用次生物质的合成提供更多底物,从而提高植物有用次生物质的含量。\n本发明提高植物有用次生物质含量的基因工程方法,包括以下步骤:1、构建嵌合基因,包括编码在植物细胞中起作用的启动子的核酸片段,和与启动子为反向连接,编码植物木质素合成关键酶CCR的核酸片段,和转录终止区;2、将嵌合基因导入当其为非转化植物时,即能在其器官或组织中产生积累有用次生物质的选自大豆或红豆杉的植物细胞中,产生转基因植物细胞;3、将转基因植物细胞生长在适宜嵌合基因表达的条件下;4、筛选与鉴别转基因植物细胞,与同类非转基因植物细胞相比,其木质素合成关键酶CCR和木质素含量被减少,而至少有一种有用次生物质的含量被提高;5、将转基因植物细胞进一步培养成为转基因植株,或转基因愈伤组织,或转基因细胞系。\n本发明所述构建的嵌合基因是反义基因结构(antisenseconstructs)。\n本发明所述构建嵌合基因,其中启动子是组成型表达启动子或器官特异表达启动子,其中组成型表达启动子为CaMV 35S,器官特异表达启动子为木质部特异表达启动子。\n本发明所述植物木质素合成关键酶为羟基肉桂酰辅酶A还原酶(cinnamoyl CoA reductase,CCR)。\n本发明所述植物当其为未转化植物时,就能在其器官或组织中积累有用次生物质,该类植物可选自大豆或红豆杉。\n本发明所述嵌合基因是通过农杆菌介导途径,电击法,微弹轰击法,显微注射法或脂质体技术导入所述植物细胞。\n本发明所述有用次生物质为由苯丙烷途径(Phenylpropanoidpathway)提供前体(底物)的产物,包括药用植物的有效成份,可具体为异黄酮或紫杉醇。\n本发明所述转基因植物细胞,含有所构建的嵌合基因,该细胞可进一步培养成为转基因植株,转基因愈伤组织或转基因细胞系。\n一种由本发明所述方法获得的有用次生物质含量被提高的转基因植物细胞,含有所述构建的嵌合基因。\n一种由本发明所述方法获得的有用次生物质含量被提高的转基因植物细胞,进一步培养形成转基因细胞系或转基因愈伤组织或转基因植株。\n本发明的有益效果是采用基因工程方法,将构建的嵌合基因导入植物细胞,并进一步培养形成为转基因植株,愈伤组织或细胞系,它们的后代能够稳定地抑制木质素合成特异酶系CCR的表达,降低其生物合成量,而使木质素生物合成的底物----芳香族氨基酸,苯丙烷途径产物更多地流入异黄酮或紫杉醇等次生物质的合成途径,提高其产量。这为提高药用植物有效成份等有用次生代谢产物的含量,提供了一条新的技术途径。\n附图说明\n图1:CCR反义植物表达载体的构建;图2:重组质粒pUCm-T-CCR的PCR鉴定P:PCR分子量标准;1:质粒pUCm-T-CCR扩增产物图3:重组质粒:pUCm-T-CCR PstI酶切鉴定M:λDNA/Hind III;E:质粒:pUCm-T-CCR用PstI酶切结果图4:质粒pACCR的PCR鉴定4-a质粒pACCR以CCPF+CCRR为引物的扩增产物P:PCR分子量标记;1:质粒pUCm-T-CCR扩增产物;\n2:质粒pIG121扩增产物;3:质粒pACCR扩增产物;4-b质粒PACCR以P35S+CCRF为引物的扩增产物。\nP:PCR分子量标记;1:质粒pIG121扩增产物;2---3:质粒pACCR扩增产物。\n图5:质粒pACCR酶切鉴定M:λDNA/Hind III;L:DNA Ladder5a:pACCR质粒SacI酶切结果。\n1:质粒pACCR;2:pACCR质粒SacI酶切;5b:pACCR质粒BamHI酶切结果。\nM:λDNA/HindIII1:pIG121质粒BamHI酶切;2:pACCR质粒BamHI酶切。\n具体实施方式\n以下举例详细说明利用本发明的实施方案,但本发明不限于下面提供的应用,以下描述不对本发明的保护范围构成任何意义上的限定。\n实施例1.木质素合成关键酶基因CCR的克隆及其反义表达载体的构建1)植物材料:供试拟南芥(Arabidopsis thaliana L.Heynh)为Columbia生态型,由中国科学院植物生理与生态研究所许政暟先生惠赠。拟南芥基因组DNA采用CTAB法从叶片中提取。\n2)菌株和质粒:克隆载体pUCm-T载体为博彩生物科技公司有限公司产品。植物表达双元载体pIG121、辅助质粒pRK2013、大肠杆菌(Escherichia coli)DH5α、HB101、根癌农杆菌(Agrobacteriumtumefacious)EHA105由浙江省农业科学院作物品质改良基因工程实验室保存。\n3)酶与化学试剂:限制性内切酶、T4 DNA连接酶、小牛肠碱性磷酸酶(CIAP)等工具酶、λDNA/HindIII购自MBI公司,DNA分子量标记PCR Marker购自华美生物技术公司,DNA Ladder购自鼎国生物技术发展中心。Taq DNA聚合酶、琼脂糖及其它生化试剂为上海生工生物工程公司产品;PCR引物由上海生工生物工程公司合成。\n4)CCR基因的扩增及克隆:根据Genbank中CCR基因序列,设计合成一对特异引物,以拟南芥基因组DNA为模板进行PCR扩增。\n5′端引物:Primer CCRF:5′-GCTGGTGGATACATCGCTTCT-3′3′端引物:Primer CCRR:5′-AGGAGAAGCCGTGTGAAAGAC-3′PCR扩增程序:扩增条件为94℃预变性5min;94℃,30s,55℃,1min,72℃,1.5min 30个循环。最后一个循环后72℃延伸10min。\nPCR扩增产物经2%琼脂糖凝胶电泳,获得一个约为343bp的扩增条带(图2),与预期大小一致。\n带单个A的PCR产物与pUCm-T载体经T4 DNA连接酶连接后,转化大肠杆菌DH5α感受态菌。用含100μg/mL氨苄青霉素的X-Gal/IPTG/LB平板筛选。小量提取阳性克隆的质粒DNA,PstI酶切后获得两条DNA电泳带,一条与载体pUCm-T的大小相近(约2.8 kb),另一条与PCR扩增片段大小相近(约343 bp)(图3),证明PCR产物已克隆到pUCm-T载体中,将此克隆命名为pUCm-T-CCR。\n5)序列测定及分析:利用位于克隆载体pUCm-T Vector多克隆位点上游的T7启动子序列,对阳性重组质粒进行自动测序。测序工作由博亚生物技术公司完成。经序列测定,克隆片段由343个核苷酸组成,其序列如下:GCTGGTGGAT ACATCGCTTC TTGGATTGTT AAGATACTTCTCGAGAGAGG TTACACAGTC AAAGGAACCG TACGGAATCCAGGTACTTGT CCATTTTTAT ATATACTTTT TTGGTCACTTGTGATAATTA TCTGAAGTTG TTGAGGTTAT TTTTATTGTTTTGATTGACA ATTTGGATTT CTTTATAATT ATTTTTCTTGCAGATGATCC GAAGAACACA CATTTGAGAG AACTAGAAGGAGGAAAGGAG AGACTGATTC TGTGCAAAGC AGATCTTCAGGACTACGAGG CTCTTAAGGC GGCGATTGAT GGTTGCGACGGCGTCTTTCA CACGGCTTCT CCT用BLAST程序在GenBank+EMBL+DDBJ+PDB中进行核苷酸序列同源性比对,结果显示该片段与拟南芥1号染色体上的CCR基因部分序列的同源性为100%。\n6)反义CCR基因植物表达双元载体的构建及鉴定:反义CCR基因植物表达结构如图1所示。为便于基因克隆,利用PCR方法在目的片段两端引入Sac I位点。以pUCm-T-CCR质粒DNA为模板进行PCR扩增,扩增产物用Sac I酶切,回收约343 bp片段;将表达载体pIG121用Sac I酶切,CIAP脱磷。载体:目的片段以1∶3的比例混合,用T4 DNA连接酶连接。转化大肠杆菌HB101感受态细胞,涂布在含100μg/mL卡那霉素的LB平板上筛选重组子。用CCRF、CCRR引物PCR筛选克隆,阳性克隆的扩增片段与对pUCm-T-CCR扩增获得的片段大小一致(图4a)。为确定目标片段与CaMV 35S启动子的反向连接,采用目标基因的5′引物CCRF和对应于CaMV 35S启动子的一段核苷酸序列为引物(P35S:5′-CGTAAGGGATGACGCACAAT-3′),进行PCR筛选,部分克隆有扩增片段,其长度略大于以P35S和GUS基因3′引物(PGUS:5′-CGCAAGACCGGCAACAGG-3′)扩增出的片段(图4b)。对该阳性克隆质粒DNA进行酶切鉴定,用Sac I酶切得到一个约349 bp的片段,与CCR基因加Sac I位点后的插入片段长度相符(图5a);用Bam HI酶切得到一个约4.1kb左右的片段(图5b),即GUS基因片段+CCR基因片段+NOS终止子片段+35S启动子片段+HPT II基因片段。PCR及酶切结果说明CCR基因片段已插入表达载体中GUS基因之后,且与35S连接方向为反义方向,将此反义CCR植物表达载体质粒命名为pACCR。\n7)反义CCR植物表达双元载体转化农杆菌:采用三亲交配法,以pRK2013为助动质粒,将反义CCR植物表达载体pACCR导入根癌农杆菌EHA105。在含有Rif(25μg/mL)和Km(100μg/mL)的YEB培养基上筛选转化体,以CCRF、CCRR引物和P35S、CCRF为引物对三亲杂交阳性克隆进行PCR扩增,并用碱法提取质粒进行酶切鉴定。酶切结果结合PCR鉴定结果表明,反义CCR植物表达载体已成功地转化了根癌农杆菌。\n实施例2.反义CCR基因转化拟南芥1)拟南芥转化采用花序喷雾法。选取饱满的拟南芥(Columbia生态型)种子,于4℃春化2~3天后播种,以16h光照/8h黑暗的光周期,于22~24℃培养,抽苔后剪去莲座叶以上主花轴,以促进次生花轴抽苔。至次生花轴2~10cm长时,用于转化。\n农杆菌的培养:挑取农杆菌EHA105(含有质粒pACCR)单菌落接种于5mL含25mg/mL Rif+100mg/mL Km的YEB培养液,200rpm,25-28℃培养过夜预活化。取50μL菌液接种于50mL同样培养液,培养至对数生长期。菌液于室温5500g离心20min。菌体用转化液(1/2MS,pH5.7含:5%蔗糖,0.02~0.05%Silwet L-77,44mMbenzylamino purine)稀释至OD600~0.80。将菌液均匀喷于拟南芥植株,以花序完全湿润为限。收集转化植株种子,在含50mg/mL Km的1/2MS平板上筛选,抗性小苗移栽到土壤中,进行分子检测。经验证的转基因植株进行木质素和次生产物含量分析。\n2)转基因植物的鉴定转基因植物的PCR检测:PCR方法可以快速地测定外源基因在转化植株或愈伤组织中的整合。由于植物本身带有内源CCR基因,因此,在用PCR或Southern方法检测转基因植株时,不能用CCR基因本身的序列来设计引物或探针。本研究采用P35S、CCRF引物来进行PCR扩增。\n提取转基因植株或愈伤组织总DNA,以此为模板,进行PCR。转基因植株扩增出了与含Anti-CCR基因的重组质粒pACCR扩增片段相同分子量的条带,而对照植株未扩增出相应条带。\n转基因植株或愈伤组织的Southern杂交分析:再生植株总DNA经限制性内切酶完全酶解后进行Southern杂交。DNA探针为超级双元载体pIG121中带潮霉素基因的片段。按TaKaRa公司随机引物标记试剂盒说明书进行标记。Southern杂交按《分子克隆》(SambrookT,TanaKa K,Monma T.Molecular Cloning:A Laboratory Manual.ColdSpring Harbor Laboratory après,New York,1989)所述方案进行。\n3)木质素含量测定:参考波钦诺克方法(波钦诺克XH。植物生物化学分析方法。1983,北京:科学出版社。165-166,178-180。)称取植物材料(鲜材料)0.2~0.5克,用1%醋酸处理分离出糖、有机酸和其它可溶性化合物。然后用丙酮处理,分离出叶绿素、脂肪和其它脂溶性化合物,再用72%(比重1。63)的硫酸分离出纤维和半纤维。离心(2500rpm),弃上清。将沉淀用蒸馏水洗涤后,在硫酸存在下,用重铬酸钾氧化水解产物中的木质素,再用碘量法测定过量的重铬酸钾。在终点前加入1mL淀粉指示剂,然后滴定到绿色。木质素含量的计算公式:x=0.433×K(a-b)DW]]>x:木质素含量(%);K:NaS2SO3的浓度;DW:样品重(g);a:对照液用去的0.5mol/L NaS2SO3体积(mL);b:滴定样品所用去的0.5mol/L NaS2SO3体积(mL)。\n4)植株酚类含量测定:参照朱广廉(朱广廉.植物生理学实验.北京大学出版社,1990:229~231)Folin-酚试剂显色法,含量以(μg/g鲜重计。\n表1.反义CCR基因对拟南芥植株木质素、酚类含量的影响\n5)纯合反义CCR转基因拟南芥植株的获得:上述反义CCR转基因植株经自交、后代经Southern、PCR等分子跟踪检测,可选育获得反义CCR基因纯合的转基因植株。\n实施例3.利用反义CCR基因提高大豆异黄酮含量\n1)植株转化:采用农杆菌介导法,以大豆子叶节为外植体,进行大豆的遗传转化,转化过程参考周思军等方法(周思军,李希臣,刘昭军等。通过农杆菌介导法将Bt(cryIA)基因导入大豆。(大豆科学,2001,20(1):157~163)。\n菌液准备:从新鲜平板中挑取单菌落,接种到含相应抗生素的YEP培养基中,27℃振荡培养至对数生长期(OD600≈0.5)。菌液在4000rpm、4℃下离心10分钟,然后将细菌重悬在1/10 B5培养基(Gamborg等,1968)中(OD600≈0.5)。重悬培养基中附加有1.7mg/LBA、0.25mg/LGA3、100μM AS(乙酰丁香酮)和3%的蔗糖,AS在高压灭菌后加入。培养基的pH在高压灭菌前调至5.4,并加入20mMMES作为pH缓冲剂。\n外植体准备:选用无病、饱满的大豆种子进行消毒;70%酒精1分钟→0.1-0.2%HgCl 15分钟→无菌水冲洗4-5次,每次至少5分钟→无菌水浸泡2小时以上。消毒后的种子接种在B5培养基(含2mg/L BA)上发芽。用发芽5-7天的无菌苗切取子叶节外植体:在离子叶节3mm左右处切去下胚轴,再切去1/3的子叶,然后在两个子叶中间将胚轴纵向切开,去掉顶芽,用解剖刀在子叶与胚轴交接处直径约3mm的范围内划5-7刀。经过上述操作,每个无菌苗可产生2个用于转化的子叶节外植体。\n转化及筛选:将制备好的子叶节外植体放入重悬后的菌液中浸泡5-30分钟。倒掉菌液,将外植体放在上下均铺有无菌滤纸的大培养皿内,吸掉多余的菌液。然后将外植体近轴面朝下接种在铺有一层无菌滤纸的共培养培养基上,共培养培养基与细菌重悬液培养基相同,加0.5%琼脂固化。培养物在24-25℃、黑暗或弱光下共培养3天后将外植体转入无菌三角瓶中,加入液体B5培养基(含有1.7mg/L BA、0.25mg/LGA3、3%蔗糖、500mg/L羧苄青霉素、20-30mg/L潮霉素,pH5.6)摇动三角瓶洗两次,或在摇床上150rpm振荡培养2-3天,每天更换新鲜培养基。然后转入固体培养,培养基成分相同,子叶近轴面朝上,下胚轴插入培养基。每两周转接一次,转接时切去下胚轴基部的老化组织。待芽可见时(约4周左右),将外植体转入芽伸长培养基。培养基成分与不定芽诱导培养基大体相同:BA浓度降为1.0mg/L或改用1.0mg/L玉米素,GA3浓度增至0.5mg/L,添加0.2mg/LIBA,加Gln和Asn各50mg/L。4-6周后再生苗可长至2.5-3cm高。切下再生苗,转入1/2 B5培养基(附加0.5-2.0mg/L IBA或NAA,2%蔗糖,0.8琼脂)诱导生根。培养条件均为22-26℃,相对湿度70-75%,光周期16/8小时。2周左右可生根。\n再生植株移栽:再生植株生根并长两片以上复叶后,将三角瓶打开盖,在气候箱中(相对湿度85%、光照强度3000Lux、温度22-26℃)锻炼5-7天。然后取出小植株,洗净根部的培养基,栽入盛有灭菌的沙、土(1∶1)混合物的小盆中,在气候箱中继续培养1-2周。在前一周时间内逐渐降低相对湿度至75%,使小植株得到逐步驯化。然后栽入大盆生长至成熟。\n2)转基因植株分子鉴定:参见实施例2。\n3)大豆中异黄酮含量的测定:采用反相高效液相法(孙梅君,骆炼,史长颖等,中国大豆制品中黄酮含量测定和分析研究,食品与发酵工业,2000,26(3)14-19)测定。\n材料:标样染料木苷和黄豆苷为Sigma产品。HPLC用甲醇、乙酸为色谱级,水为高纯水,其余试剂为分析纯。\n主要仪器:高效液相色谱仪(日本岛津LC-9A)。\n色谱柱:Shim-pack clc-ods C-18 0.15m×6.0mmФ;紫外检测器:SPD-6AV;流动相:甲醇∶5%乙酸=30∶70;流速:1.0mL/mim(8.5min前),1.5mL/min(8.5min后);波长:224nm;检测灵敏度:0.08AUFS;柱温:50℃。\n将大豆种子磨碎后过40目筛,然后用正已烷脱脂,用80%甲醇回流萃取,萃取液经0.45μm微滤后,取10μL进样,经紫外检测器检测,通过与标样比较,根据标样的保留时间定性,根据标样的峰面积定量。\n表2.反义CCR基因对大豆种子异黄酮含量的影响\n4)纯合反义CCR转基因大豆植株的获得:上述反义CCR转基因植株经自交、后代经Southern、PCR等分子跟踪检测,可选育获得反义CCR基因纯合的转基因大豆品系。\n实施例4.利用反义CCR基因提高红豆杉愈伤组织紫杉醇含量1)反义CCR转基因红豆杉愈伤组织的获得:采用基因枪转化法。\n外植体的消毒:选择1~2年生具有绿皮处于阴暗位置的南方红豆杉嫩枝作为供试材料,枝条洗净后剪成小段,分别用70%乙醇和0.1%HgCl2消毒2min和20min。用无菌水漂洗6次后,将茎段剪成长约1.5-2.0cm的小段,斜插在诱导培养基上。愈伤组织诱导培养基为B5培养基的无机盐,添加100.0mg/l肌醇、1.25mg/L烟酸、1.0mg/L维生素B1、0.5mg/L维生素B6、20.0g/L蔗糖、1.0g/L水解乳蛋白、0.1mg/L BAP、2.0mg/L 2,4-D、琼脂8.0g/L,培养基pH值为5.8。在黑暗培养室中培养。培养温度为22±2℃。约20天左右出现愈伤。诱导结束时,将外植体上形成的较大块愈伤组织(直径0.5cm以上)剥离下来,接种在增殖培养基上继代培养。愈伤组织增殖培养基同诱导培养基,但无水解乳蛋白,2,4-D浓度为1.0mg/L。在光照培养室中培养,培养温度为22±2℃,光照强度约为1500 lx,光周期为14h。\n经1-2次继代的愈伤组织用于基因枪转化。愈伤组织于轰击前24h,转移于高糖培养基上(含蔗糖5%,其余成分同增殖培养基)。微弹DNA制备:质粒pCCRA采用碱法提取,质粒DNA的浓度测定按其OD260值进行估算,用TE调整到浓度为1μg/μL。所用金粉颗粒直径为1.6μm。无菌条件下制备金粉的乙醇溶液,浓度60μg/μL。取50μL悬浮液分装于Eppendorf管中,每一管加入50μg质粒,50μL2.5μgCaCl2和20μL 1.0M亚精胺溶液。充分振荡3min,10000rpm离心10min,弃上清,沉淀重新悬浮于50μL无水乙醇中。每次轰击用10μL。\n基因枪轰击:使用Bio-Rad公司的PDS-1000/He型气动式基因枪进行转化,按产品说明书进行操作。真空度26-30英寸汞柱,轰击压为1100psi,每皿轰击一枪。\n轰击后愈伤材料于高糖培养基上培养过夜后,转移到无抗生素的固体增殖培养基上,培养4~6d后,转移至含潮霉素(20mg/L)的抗性增殖培养基上,每4周转移1次,选择生长正常的抗性组织。\n2)转基因愈伤组织分子鉴定:参见实施例2。\n3)红豆杉愈伤组织紫杉醇含量的测定:采用高效液相色谱法(孙珺,胡正海。红豆杉愈伤组织的培养及紫杉醇的产生。西北大学学报。2000,30(1)55~59)检测红豆杉培养物紫杉醇含量。\n仪器和试剂:日本岛津公司LC-9A型高效液相色谱仪;Shim PackCLC-ODS色谱柱;CR-9A机记录、计算。甲醇、三氯甲烷为分析纯。HPLC用甲醇、乙腈为液相色谱洗脱剂;水为高纯水;紫杉醇及三尖杉酯碱标准品均为Sigma公司产品;流动相为甲醇-乙腈-水(25∶24∶41);流速1mL/min;检测波长为227nm。\n标准曲线的制作:精确称取紫杉醇标准品,配成1mg/mL的甲醇溶液。再准确吸取标准品溶液2.00,4.00,6.00,8.00,10.00μL注入液相色谱仪,记录其色谱图,分别量取峰面积A,重复3次,取其平均值。以峰面积A为横坐标,进样量m为纵坐标作图得一直线,即为标准曲线。从该曲线可得出回归方程:m=bA+c,其中b,c为常数,方程的线性相关系数应为r=0.999~1.000。\n样品处理及紫杉醇含量测定:将培养30d后的愈伤组织在60℃下烘干至恒重,研碎至0.25mm。以氯仿∶甲醇=6∶4回流提取3次,浓缩、石油醚脱酯,然后减压真空低温干燥至恒重。\n准确称取待测样品适量,加流动相溶解,配成浓度为0.3mg/mL的待测溶液,精密进样分析。重复3次,测得峰面积Ai,取其平均值,代入回归方程,求出mi。百分含量Pi%按下式计算:Pi-=mi/mw×100%,其中mw为样品质量。\n表3.反义CCR基因对红豆杉愈伤组织紫杉醇含量的影响\n实施例5.利用反义CCR基因提高红豆杉细胞系紫杉醇含量1)红豆杉愈伤组织的诱导:参见实施例4。\n2)红豆杉悬浮细胞系的获得:对诱导发生的红豆杉愈伤组织培养物进行比较、选择,初步检测培养物中紫杉醇含量。获得生长较快、紫杉醇含量高的无性系组织用作后续研究。\n将优良无性系愈伤组织材料接种到B5(含有2,4-D1.0mg/L,IAA1.0mg/L,KT 0.1mg/L)培养基中进行悬浮培养,每250mL三角瓶分装60mL左右的液体培养液,每个三角瓶接种量约150mg(干重)。摇床转速120r/min,其他培养条件同固体培养。两周后,在无菌条件下先用筛网滤去较大颗粒,再滤取细小细胞团。选取3-5个细胞聚集的小细胞团,散开接种到培养皿的琼脂培养基上进行培养。20d左右,待小细胞团长大后,挑选色泽、形态和生长状态不同的细胞团,分别切割并接种到50ml的小三角瓶中进行培养。选择生长良好的细胞团做进一步的观察与选择。将选为进一步观察比较的细胞团平分为二:一部分继续培养并做为选择材料;另一部分扩大培养并做含量检测。经观察与选择,将生长慢、紫杉醇含量低的细胞团从选择培养的材料中剔除;将余下的细胞团再分割成小块,并接种到新的培养基上,作新一轮的观察、比较、检测。经多轮选择,最终选择出紫杉醇含量高、生长良好的细胞系材料,将其进行扩大繁殖和继代培养,并作进一步去杂保纯,待其培养材料性状稳定后,作为优良细胞系用于培养试验与扩大培养。\n3)反义CCR基因转化红豆杉悬浮细胞:采用基因枪转化,微弹DNA制备、基因枪轰击条件参见实施例4。转化后红豆杉悬浮细胞转移至含至含潮霉素(20mg/L)的液体培养基筛选,同法培养未转化红豆杉悬浮细胞做对照。对筛选后存活的细胞扩大培养,并进行PCR检测。\n4)红豆杉悬浮细胞紫杉醇含量的测定:收获待测的培养物,在60℃下烘干至恒重,研细,过40目筛。采用高压液相法测定其紫杉醇含量,方法参见实施例4。\n表4.反义CCR基因对红豆杉悬浮细胞系紫杉醇含量的影响\n以上以举例方式详细描述了本发明的实施过程,但对本领域熟练技术人员来说,显而易见的是,在实施过程中可进行许多等效的修改和替换。因此,本发明的范围仅以权利要求书的限定为准。
法律信息
- 2012-10-03
未缴年费专利权终止
IPC(主分类): C12N 5/10
专利号: ZL 03142134.2
申请日: 2003.08.08
授权公告日: 2007.05.30
- 2007-05-30
- 2005-02-09
- 2004-07-21
引用专利(该专利引用了哪些专利)
序号 | 公开(公告)号 | 公开(公告)日 | 申请日 | 专利名称 | 申请人 |
1
| |
2002-01-02
|
1999-10-06
| | |
被引用专利(该专利被哪些专利引用)
序号 | 公开(公告)号 | 公开(公告)日 | 申请日 | 专利名称 | 申请人 | 该专利没有被任何外部专利所引用! |